

In the book Metazoa, I connect the evolution of subjective experience (consciousness, in a very broad sense) to some ideas about nervous system activity. The main idea I look at is the possible importance of large-scale rhythmic patterns, some of which can be detected with the “EEG” technology. I also look at the more controversial idea that electromagnetic fields themselves, created by nervous system activity, might have a special role in the explanation of experience.
Starting with the rhythms: this aspect of brain activity was uncovered especially by Hans Berger in the 1920s, as part of a search for ESP. Berger had predecessors in his study of electrical activity in the brain, but he was the one who invented the EEG technology and made recordings with it. His technology is special because measurements can be done on the scalp itself, and can detect electrical rhythms with fine-grained temporal resolution.
On the right is an EEG readout done by Berger himself. 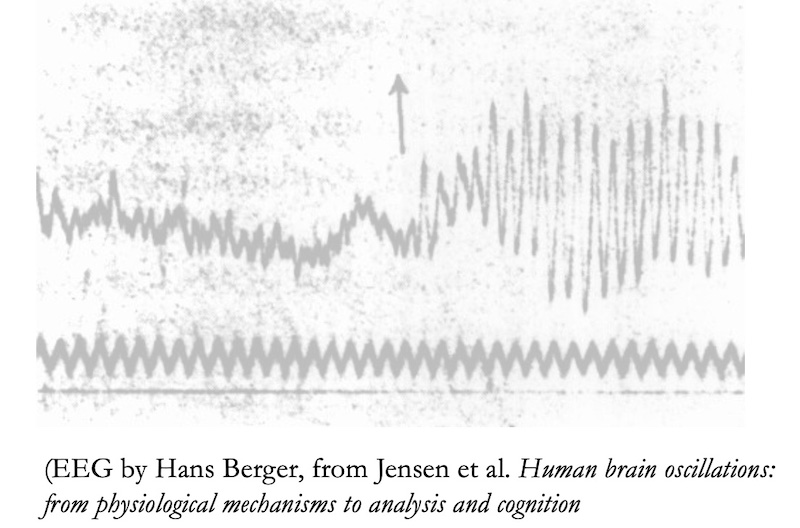
The color diagram is from the Mayo Clinic.
There’s now quite a long history of claiming that the kind of rhythms that EEG methods detect are important to how our brains work, and specifically important to felt experience or consciousness. The idea seemed to become important round the late 1980s (eg., Rodolfo Llinás) though I don’t yet know the details of the history. It was brought to a wider audience by Francis Crick and Christof Koch in their 1990 paper about “neural correlates of consciousness.”
What might these electrical rhythmic do? They might figure in integration, unification, organization – those term suggest somewhat different things but are closely related. One view could be expressed as follows: nervous system activity includes networked, cell-to-cell influences that operate through synapses and action potentials (spikes), and also includes more diffuse, large-scale forms of electrical activity that have an organizing or unifying role. That’s the idea (or one idea) I am interested in.
I am not a neuroscientist, and a lot of neuroscientists doubt that anything like this is true. Plenty of people think that these brain rhythms are just byproducts – the hum of the machine – and don’t make anything, or much of anything, happen. One reason I gave them so much attention in Metazoa is an “in principle” point. If these phenomena – which are straightforwardly physical – were important, it would make available a different view of how neural activity can be a basis for experience. The problem would look different from how it has often appeared.
That would be interesting even if the brain does not actually work in the way I was exploring. It’s much more important, of course, if the brain does work like that.
Over just the last couple of years, the empirical fortunes of these ideas about the importance of large-scale dynamic patterns seem to have risen. They’re still oscillating – appropriately – but seem to me to be gaining more support. I am giving talks in this area at the moment and writing new papers. To help with this process, I am going to write a series of long posts here that work through details, note disagreements and corrections to things I’ve said, link these ideas to more radical ones, and so on. These posts won’t be at all consistent in their accessibility; sometimes I’ll go through simple things slowly, sometimes quickly add something much more complicated (at least for me). This is the first of at least three or four posts. Each will have photos mixed in that don’t have a strong relationship to the ideas in the text – often just photos that belong somewhere on the site. The photos in this post are extra octopus photos from a dive earlier this year at the Pipeline in Nelson Bay.

_________
Let’s start with EEG measurements themselves. Electrodes on the scalp pick up rhythmic electrical patterns. What are they picking up? To my surprise, there still seems to be some uncertainty about this – neuroscientists certainly say different things about it. This much is clear: the EEG recording is tracking ion flows (flows of charged particles) within the brain. These are ion flows across cell membranes. What kinds of ion flows? The obvious candidates are “action potentials” or “spikes,” the electrical fits that neurons go through (also referred to as neurons “firing”). These are short and dramatic events in which positively charged ions rush into a neuron, changing the electrical properties of the whole cell, before the normal state is restored. Below in this post I have a recent quote from a neuroscientist saying that large numbers of action potentials are the basis for EEG recordings. When I discuss large-scale dynamic patterns in talks, sometimes someone in the audience says that this is not something distinct from the familiar cell-to-cell network interactions; it’s just a lot of them measured in a coarse-grained way. It’s important, at least to some exent, to the idea that large-scale rhythms have a special role in the mind that this is not true, and I take the mainstream view within neuroscience to be that this is not true – the EEG is not just a large-scale read-out of spikes. Instead, an EEG is reading a to-and-fro of ions across membranes that is below the threshold that sets off a spike – a fainter and slower flow. In these posts I’ll often use György Buzsáki’s 2006 book Rhythms of the Brain. This is how he describes it:
[W]e distinguish excitatory postsynaptic potentials (EPSPs; or currents, EPSCs) from inhibitory postsynaptic potentials (IPSPs; or currents, IPSCs). Compared to the fast action potentials, membrane potential changes associated with EPSPs and IPSPs are several-fold smaller in amplitude. However, they last for tens of milliseconds. This latter property is critical for understanding the generation of EEG activity.
[T]he contribution of action potentials to the local field and especially to the scalp EEG is negligible.
In this view, there is constant input into neurons in the form of ion flows. This input can cause a spike, but it need not. The basis for the EEG is rhythmic ionic inputs of this kind. Buzsáki I take to be a leading authority in this area. Importantly, though, in his 2006 book he distinguishes between what he calls a “classical” picture on this topic and some newer ideas. This is a footnote:
- This “classical” description of the origin of extracellular fields must be supplemented by the recent findings about the active properties of neurons (see Cycle 8; Llinás, 1988). Subthreshold oscillations, afterpotentials, Ca2+ spikes, and other intrinsic events also produce relatively long-lasting transmembrane events. The contribution of these nonsynaptic events to the local field potential can often be more important than the contribution of synaptic events (Buzsáki et al., 2003b).
When I say the footnote is “on this topic,” I need to be explicit. This note is about the generation of electric fields surrounding neurons in general. Those are the basis for an EEG recording, and also the basis for something related, an LFP (local field potential) recording. An LFP recording is taken using an electrode interted into the brain, not sensors on the scalp. LFP recordings can be done with many more animals than an EEG can, as use of an EEG is dependent on a head and brain architecture of a certain kind. I take Buzsáki to be saying something here about the basis for EEG recordings and the basis for LFP (and MEG) recordings – all of them. I am going to chase down the references Buzsáki gives there in another post. For now, the main ideas are: (i) Sometimes people say the EEG is just recording spikes, but Buzsáki and others say this can’t be true; (ii) It is more accurate to summarize things by saying the EEG recording is picking up spatially organized flows of ions that are inputs to neurons at synapses; (iii) Other phenomena also play a role (“Subthreshold oscillations, afterpotentials…”).

_________
EEG recordings are the tip of an iceberg. Or rather, the basis in the brain for EEG recordings is the tip of an iceberg. We can get EEG recordings because the neurons in a human cortex have a particular spatial arrangment, in parallel, and it must help that these cortical neurons are also close to the skull, on top of the brain. This creates a signal so strong that it can be picked up from outside the brain using 1920s technology. As my discussion of that footnote in Buzsáki’s book indicates, there’s a lot more going on, buried inside our brain and also in brains that can’t support EEG recordings. The basis for the EEG recording is some especially strong and well-organized electrical rhythms – the tip of what might be a biologically and philosophically important iceberg. I mentioned LFP methods above, which pick up more “local” electrical oscillations in a specific part of the brain. There is also the MEG technology – magnetoencephalography, which detects magnetic rather than electric field activity. Whenever there’s a flow of electric charge, a current, a magnetic field is created as well. Changes in these fields can also be picked up from outside the brain, but with a much more complex device than an EEG. The machine, aptly, is called a SQUID (superconducting quantum interference device).
In the book Metazoa, I introduced this topic involving rhythms and fields in a chapter about fish, and I also noted the interest that Berger (the EEG inventor) had in ESP. In the first published versions of the book, I have a sentence that is a bad mistake. I wondered about the transmission of electro-rhythmic influences from one fish to another, and said that this would be more feasible with fish than with humans, because air is a bad electrical conductor whereas water is a good one. I said that an “electric field” requires a conductor of the right kind (p. 92). This is not true – an electromagnetic field can propagate through air or a vacuum (or else we could not see the stars, for a start). What I said is true of currents, but not of fields. I don’t know how I came to write that (and it’s not an error introduced by an editor, or anything like that). A phenomenon I’ve seen in writing of this kind is that sometimes an author, myself or someone else, simplifies some some tricky material over and over to make it accessible, and ends up writing a version that includes a mistake. Most versions of Metazoa don’t have the mistake (paperback, e-book, most translations) but some do, and I wish they did not.
For the rest of this post, I’ll explore some details relating to the material above, give some quotes and references, and so on. In the second post I’ll start looking at the relations between rhythms across different animals – exploring other parts of the iceberg.
_________
Details
1. The relationship between spikes and EEG readings: this is from R. Douglas Fields’s book Electric Brain (p. 78). There does seem to be real disagreement in this area (I doubt that Fields is deliberately simplifying(?)).
[R]ecording the impulses of a single neuron is a slow, difficult, and tedious process. It requires delicate brain surgery, carefully maintaining the respiration and body temperature in the animal under anesthesia, as well as the ability to extract information and efficiently perform an experiment in the heat of the moment. In contrast to the painstaking process of this “single neuron” recording, the combined activity of large groups of neurons firing together causes fluctuations in the surrounding electric field that can be detected by electrodes placed on the scalp. Yes – we have returned to brainwaves. The activity recorded from outside the skull by an electroencephalogram (EEG) can be thought of as a bit like the roar of a stadium heard from the parking lot.
2. Buzsáki on the chemicals
Buzsáki’s Rhythms of the Brain is a multi-faceted book (and one not written at a consistent level). Mixed in with his exposition of the basic features of brain activity are reflections on the brain’s chemistry and its origins. A resonant passage (p. 86):
Like virtually any cell in the body, neurons have a high concentration of ions of potassium (K+) and chloride (Cl–) inside and keep the sodium (Na+) and calcium (Ca2+) ions outside. This arrangement produces a small battery that maintains a voltage difference of –60 millivolts relative to the world outside of the cell membrane. This ion separation is perhaps attributable to our single-cell ancestors and where they came from: the sea. Given the high concentration of Na+ in sea-water, keeping Na+ outside the cell was a smart choice. However, when more developed organisms migrated to land, they had to carry the sea with them to maintain the same extracellular environment. For this purpose, the circulation of lymph and blood developed. All our cells are constantly bathed in water, more precisely, salt water. [Nb: I can’t find a way to superscript those “+” symbols for charge, etc.]
Why was it a “smart choice” to keep the sodium outside? I am not sure if he’s saying that there’s just too much sodium around in the sea for cells to work well if they routinely have the same sort of level inside them, or whether he means that sodium ions, which are plentiful in the sea, provide a good device for signaling and control, as they can rush in (following a concentration gradient) and change the electrical properties of the cell quickly. Anyway, once they’re inside, something has to be done to restore both the electrical properties of the cell (the battery-like feature) and the chemical properties.
Pumping all the excess Na+ out of the neuron is a lengthy process. To regain the resting voltage across the membrane more rapidly, neurons opted for another strategy: voltage dependence of K+ channel activity. As the action potential reaches its peak, the voltage-dependent K+ channels are activated and quickly repolarize the cell. This fast repolarization is the falling phase of the action potential…. Thus, the positive charge created by the influx of Na+ is compensated for by the quick efflux of equal charges carried by K+.
An outflow of potassium is used to restore the normal electrical setting, while a slower process cleans up the chemistry. Buzsáki, like many people, emphasizes how slow this is in comparison to a computer – the sodium-potassium “push-pull” (as he calls it) takes about a millisecond, a thousandth of a second. I never get over how fast that is. The cell then takes a few more milliseconds to recover (the exact time depends on the cell). The fastest observed rate of firing by human neurons seems to be around 400-500 times per second.
The picture he offers is one where even though action potentials are, or might be, central to neural activity, there is a lot of electro-chemical back-and-forth going on at different scales that makes this possible. A variety of ion flows move continually across membranes in a slower and more “analog” way. EEG and LFP measurements are sensitive to these flows.
The harder version (p. 89):
The local field potential (i.e., local mean field), recorded at any given site in or outside the brain, reflects the linear sum of numerous overlapping fields generated by current sources (current from the intracellular space to the extracellular space) and sinks (current from the extracellular space to the intracellular space) distributed along multiple cells…. The low resistance or “shunting” effect of the extracellular fluid, the membranes of neurons, glia, and blood vessels, and the slow movement of ions attenuate current propagation in the extraneuronal space. Because the passive neuron acts as a capacitive low-pass filter, this attenuation is quite discriminative: it affects fast-rising events, such as the extracellular spikes, much more effectively than slowly undulating voltages. As a result, the effects of postsynaptic potentials can propagate much farther in the extracellular space than can spikes. Furthermore, because of their longer duration, EPSPs and IPSPs have a much higher chance to occur in a temporally overlapping manner than do the very brief action potentials. Finally, EPSPs and IPSPs are displayed by many more neurons than are spikes because only a very small minority of neurons reach the spike threshold at any instant in time. For these reasons, the contribution of action potentials to the local field and especially to the scalp EEG is negligible.
Time for a break.
The photo below is from the Mediterranean coast of France, where I wrote most of this post.

If you write a comment on the post from this front page, the letters will look very small as you type. Write from here to avoid the problem.