
This is the second in a series of posts about brains, rhythms, fields, and consciousness. Some of the photos in this post are from a visit to Heidelberg University in May for a workshop where I gave a talk on these themes.Their Centre for Organismal Studies has, in its lecture room area, an elaborate physical display illustrating evolution, with model animals arranged in a large phylogenetic tree.

I assume these are not taxidermied animals, but wholly synthetic. Australian animals are especially well represented.

I like this modern style of highly accurate synthetic model-animal building. The old taxidermied style seen in museums can be magnificent, if excessively poignant, but the animals are often getting threadbare now. The new artificial ones can have a real vivacity about them.
Onward into the rhythmic brain.
Two topics in this post. First, I’ll work through the start of a paper about the physical basis for EEG readings, with the good subtitle “A review for the rest of us,” by Alice Jackson and Donald Bolger.
This is, again, just an account of the conspicuous rhythms picked up in an EEG, not an account of all brain oscillations. In the previous post, I said that the brain rhythms seen in an EEG are “the tip of an iceberg.” The “for the rest of us” review gives a strong sense of this, and of how lucky (perhaps, and in a very specific sense of ‘luck’) it is that they exist in their conspicuous form.
Their first point: the “primary source of the EEG signal” is “synchronized synaptic activity in populations of cortical neurons (pyramidal cells organized along cortical columns).” A particular kind of cell is organized in parallel columns, with the top of these cells not far from the skull. The main “input” end of these neurons is at the top. When these cells are excited (without this necessarily causing an action potential) by other cells, the flow of positive ions into the cell changes the electrical properties not only of the cell, but of its local environment.
When an EPSP is triggered at a dendritic synapse, the local extracellular fluid becomes more negative with respect to the intracellular fluid as depolarizing current [positive ions] flows into the neuron. That current will flow out elsewhere along the neuron, which produces a local, albeit more distributed, extracellular positivity.
An EPSP is “excitatory” input to the cell, making its interior more positive than before and the exterior more negative. There can also be “inhibitory” inputs to these neurons, making the interior more negative and exterior more positive. Then “volume conduction,” a current moving in a pool (rather than through a wire or across a membrane) leads to these effects propagating within the brain. Finally, we get to a picture like this. The wire at the top is from an electrode stuck on the skull.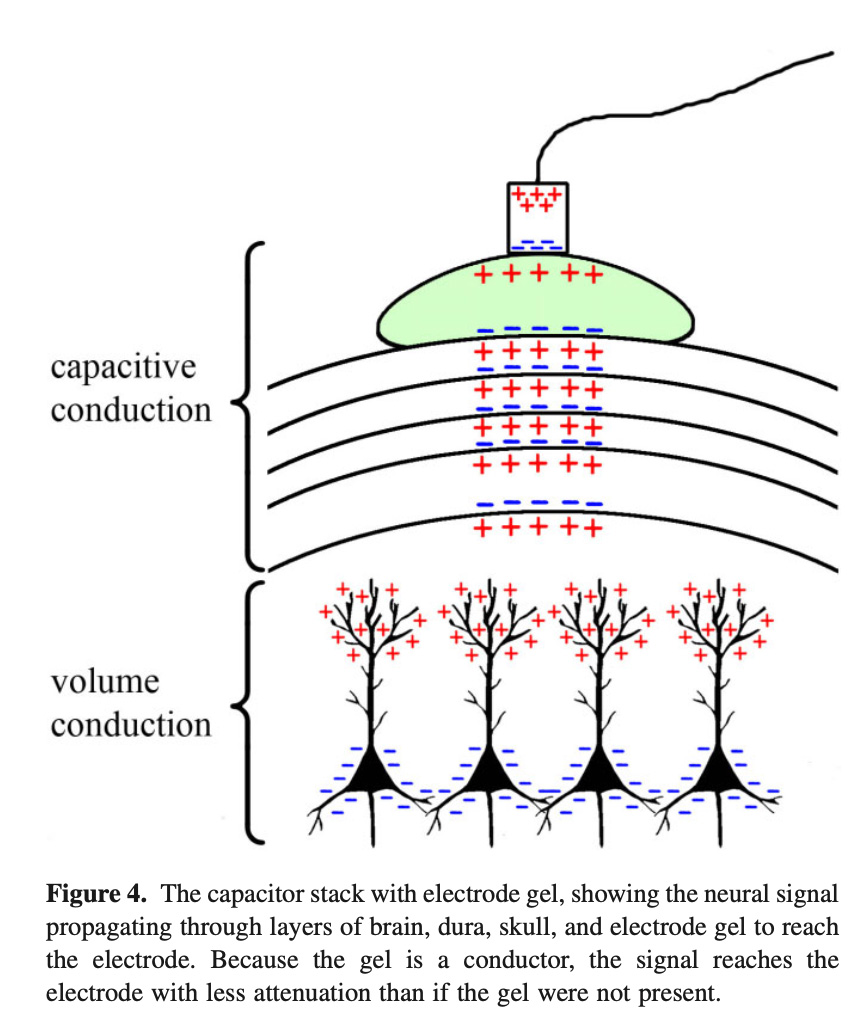
What we’re seeing: parallel neurons at the bottom are synchronized in their activity and receiving (in this case) inhibitory inputs. The fluid around their tops (dendritic trees) becomes more positive. The volume conduction hits an insulator, an obstacle. On the other side of the insulator is another conductor. The layers act as a capacitor, with charge of one kind accumulating on one side of the insulator and an opposite charge accumulating on the other side.
A capacitor is, in its most general form, two pools of charges separated by an insulating layer (a dielectric)….[I]f an insulating layer is present, then a charge difference may build up across the insulating layer, wherein negative ions push up against one side of the membrane and positive ions accumulate on the other side…. The sequence of layers from the brain to the dura layers, skull layers, scalp layers, electrode gel, and electrode thus forms a series of conductive volumes separated by insulating layers—akin to a stack of capacitors.
A detailed explanation of the figure above:
Consider an IPSP arriving at a synapse near a soma in cortical layer IV. The extracellular positivity will propagate via volume conduction to the near side of the closest insulating layer, at which point volume conduction cannot carry the signal further because the insulating layer is a physical barrier to ion flow. The positive ions accumulated at the near side of the insulating layer will repel positive charges on the other side of the insulating layer. The positive charges repelled and thus accumulated on the far side of the insulating layer will, in turn, push other positive charges away. This process will repeat through every capacitor in the stack until it reaches the electrode.
The electrode is at the very top:
An electrode’s output is a single voltage per measurement typically taken every 1–2 ms (or 1–0.5 kHz). This measured voltage reflects the sum of all charged ions that are, through the capacitive stack discussed above, pushing or pulling on the electrons in the electrode at the time that the measurement is taken. A measurement of voltage at any position on the scalp will consist of the sum of influences from many sources of electric fields because the brain is full of dipoles.
That, according to the review, is the causal path from neuron to EEG. This is along the same lines as what Buzsáki called the “classical” view of the source of EEG measurements, discussed in the previous post. And, again, all this is just part of the oscillatory story. It’s how a signal strong enough to show up outside a skull in a way detectable using 1920s technology comes to exist.

Theme 2 of this post: In the Buzsáki passages from the previous post, he distinguishes the “classical” view of the source of EEG from some other ideas, citing a 1988 paper by Rodolfo Llinás along with one of his own. That Llinás paper appeared in Science, includes a detailed review of empirical results, but also offers some adventurous, general, telegraphically-expressed ideas about how brains work. The paper is here.
First, if we go back to the outline of the EEG source above, why does any of this 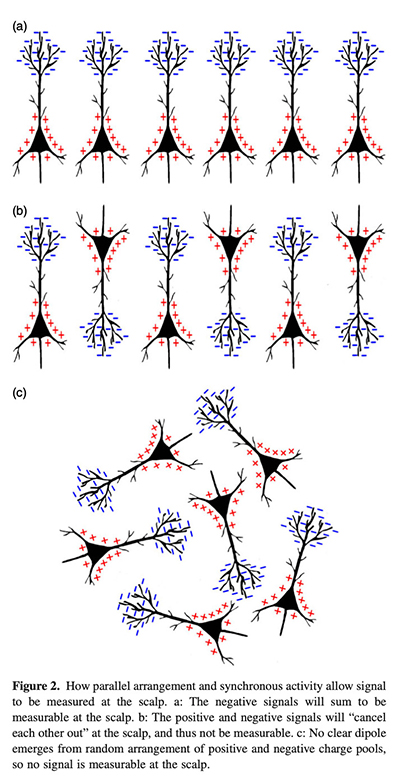 happen at all? On the classical view, I take it, it happens because neurons are influencing each other with signaling molecules, and the main way a neuron is influenced is by prompting ion channels to let flows of ions in and out of the cell. An action potential is a large-scale effect of this kind, and smaller-scale effects are ones in which one cell prompts another to let some ions in or out in a “fainter and slower” process than an action potential, and that might help lead to an action potential. Ions are induced by signaling molecules to flow into cells especially at the dendrites; the excess ions have to be sent back to the outside of the cell (at sites some way from the dendrites) in a “return” current. And if this process is both spatially and temporally organized across many cells (see the figure from Jackson and Bolger on the right), there’s something that the EEG picks up. The existence of rhythms in this system would have to be due to some rhythmic patterning in the inputs to the neurons – patterning in the way neurons are affecting others using signaling molecules. Otherwise, why would there be a rhythm as opposed to some quite different pattern?
happen at all? On the classical view, I take it, it happens because neurons are influencing each other with signaling molecules, and the main way a neuron is influenced is by prompting ion channels to let flows of ions in and out of the cell. An action potential is a large-scale effect of this kind, and smaller-scale effects are ones in which one cell prompts another to let some ions in or out in a “fainter and slower” process than an action potential, and that might help lead to an action potential. Ions are induced by signaling molecules to flow into cells especially at the dendrites; the excess ions have to be sent back to the outside of the cell (at sites some way from the dendrites) in a “return” current. And if this process is both spatially and temporally organized across many cells (see the figure from Jackson and Bolger on the right), there’s something that the EEG picks up. The existence of rhythms in this system would have to be due to some rhythmic patterning in the inputs to the neurons – patterning in the way neurons are affecting others using signaling molecules. Otherwise, why would there be a rhythm as opposed to some quite different pattern?
The part I am emphasizing here is the way the causal chain starts with signaling molecules at synapses, which prompt the EPSPs and IPSPs described above. These would have to be rhythmic in order to give rise to an EEG rhythm. The Llinás paper adds to, or changes, all this.
Llinás says that for a time (mid C2o?), “complexity in the mammalian brain was believed to be attained by the connectivity of close to ideal or “Platonic” nerve cells serving as threshold elements.” Neurons received input, and either fired or didn’t. Work on invertebrates began to show more complexity in the electrical behavior of individual neurons, but it took a while for this to be thought relevant to mammals (which are Llinás’s concern). Eventually, extra complexity was uncovered, and this includes (relevant for us) the fact that:
[S]ome central neurons have ionic conductances organized to endow them with electrical autorhythmicity. In addition, in many neurons the kinetics of these ionic voltage-dependent conductances are such that the cells may respond preferentially to inputs at a certain frequency or frequencies, that is, they behave as resonators.
He discusses examples. And then begins a section with: “Two Main Forms of Brain Activity”
How do the oscillatory properties of central neurons relate to the information-carrying properties of the brain as a whole? In principle, one may propose that intrinsic electroresponsiveness generates internal computational states that serve as the reference frame, or context, for incoming information. That is, brain function is proposed to have two distinct components, one which is private or “closed” and is responsible for qualities such as subjectivity and semantics, and an “open” component that is responsible for sensory-motor transformations that deal with the relations between the private component and the external world.
This is in the vicinity of the two-activity views that I said (in the previous post) that I am interested in. (Me: “nervous system activity includes networked, cell-to-cell influences that operate through synapses and action potentials (spikes), and also includes more diffuse, large-scale forms of electrical activity that have an organizing or unifying role.”) In Llinás’s version, there are “intrinsic” patterns of activity that have an organizing role (“reference frame, or context”) and an “open” component that is responsive to the external world. It’s a somewhat different distinction, but related. He adds, without argument here, that the first component is responsible for “subjectivity and semantics” – he makes an immediate mapping between his forms of brain activity and questions about experience or consciousness. Working within his distinction, I’d say that the combination is likely responsible for “subjectivity and semantics,” not one component. “Semantics” involves reference to external conditions, and “subjectivity” relates to point of view, perspective, and that will involve both sides. But there I’m guessing about how I’d develop a view in Llinás’s terms.

Llinás goes on:
More precisely, the intrinsic activity is proposed to be part of the vectorial coordinate space that allows sensory-motor transformation to occur in the context of the particular functional state of the CNS at a given instant (39). For instance, attention or expectation (both intrinsic functional states) can modify the relevance of given sensory stimuli.
This relates to a detailed and specific hypothesis about geometry and brain function that Llinás developed with Andras Pellionisz. I’m not going to try to understand that next step (and don’t know how it’s regarded now). An interesting feature of the 1988 Llinás paper is the idea that neurons, individually, have oscillatory tendencies that might be raw materials for cognitively significant forms of brain activity, and that early statement of a breakdown of roles – two forms of brain activity – with an attempt to link the distinction to subjectivity.
Returning to the connections between the first and second parts of this post: on the “classical” view, neural oscillations that show up in an EEG must come about because of patterning in the signaling inputs that neurons receive (it’s EPSPs and IPSPs that are being tracked). Llinás is saying that neurons don’t have to be “told” to oscillate; some of them will do so as a result of their intrinsic electrical activity. So far, that’s just something at the level of individual cells; there’s no reason yet to expect large-scale synchronization. (Yet….)

In the next post I will work through yet another general account of electrical oscillations in the brain, by Buzsáki, Anastassiou, and Koch. This one introduces another presently controversial factor: ephaptic coupling. This “one-person journal club” exercise is turning out to be quite helpful for my thinking, and I might continue it for quite a while.
Other Notes
1. Llinás is an important figure in this area. I didn’t know the following: “He received his PhD in 1965 from the Australian National University working under Sir John Eccles.” And “IPSPs were first investigated in motorneurons by David P. C. Lloyd, John Eccles and Rodolfo Llinás in the 1950s and 1960s.” Both via Wikipedia. I worked at the ANU for a while; did not know this. Themes here will return when I write something I’m planning with a very different role.
2. “Specific behavioral manipulations can produce selective changes in particular brain areas. Raising kittens and mice for 3-4 months in complete darkness seriously affects not only their visual abilities but also their sleep oscillations.” From the Buzsáki book. Do not do this. I’ll write something else soon-ish that has been prompted by encountering so many cat and kitten experiments in my attempt to understand these aspects of brain functioning.
3. Second-last photo is a Thylacine, an extinct Australian tiger-like marsupial.
If you write a comment on the post from this front page, the letters will look very small as you type. Write from here to avoid the problem.